

Các dạng tổn thương DNA và phương thức sửa chữa tổn thương DNA
Các dạng tổn thương DNA
Sau khi chiếu xạ UV, giữa hai thymine và cytosine gần nhau có thể hình thành chuỗi butyrylcholinesterase dạng vòng bằng khớp nối đồng hóa trị, cấu trúc này được gọi là dimer pyrimidine. Sự hình thành dimer chính là hình thức gây tổn thương chính của UV đối với phân tử DNA.
Hình 1: Sự hình thành dimer

Hình 2: Cấu trúc của chuỗi butyrylcholinesterase

Sau khi tia X, tia y chiếu vào tế bào, gốc tự do được sinh ra từ nước trong tế bào vừa có thể làm liên kết hydro giữa các sợi kép trong phân tử DNA gẫy ra, vừa có thể khiến sợi đơn hoặc sợi kép của nó gẫy ra. Alkyl hóa bleomycin và methanesulfonate có thể gây ra đứt chuỗi. Mitomycin C có thể tạo thành liên kết ngang giữa chuỗi đơn của phân tử DNA, điều này thường xảy ra tại vị trí giữa các guanine của hai đường chéo nhau của chuỗi đơn. Liên kết ngang của chuỗi cũng thường dẫn đến hiện tượng gẫy phân tử DNA.
Phân tử DNA còn có thể xảy ra thay đổi codon cá biệt hoặc nucleotit. Tác nhân gây tổn thương DNA còn có thể cùng lúc gây nên mấy dạng tổn thương, kích thước và loại hình tổn thương liên quan đến tác nhân và trạng thái chu kỳ tế bào.
Phương thức sửa chữa tổn thương DNA
a ,Quang phục hoạt (Photoreactivation)
Quang phục hoạt (photoreactivation) hay sửa sai nhờ ánh sáng (light repair). Sau khi xử lý tia tử ngoại gây đột biến, nếu đưa ra ánh sáng thì phần lớn sai hỏng được phục hồi nhờ enzyme photolyase. Enzyme này gắn vào photodimer cắt nó thành các monomer dưới tác dụng của ánh sáng mặt trời có bước sóng 320-370 nm. Sau đó phục hồi các base ban đầu. Quang phục hoạt được phát hiện ở vi khuẩn, men, động vật nguyên sinh, loài rong, ếch, chim, động vật có túi và lympho và tế bào nguyên bào sợi. phương thức sửa chữa này rất phổ biến nhưng chủ yếu có ở sinh vật bậc thấp. Cùng với sự tiến hóa của sinh vật, tác dụng của phương thức này cũng ngày càng ít đi.
b, Sửa chữa bằng cắt bỏ (excision repair)
Phần lớn các cơ chế sửa sai khác thực hiện theo lối cắt bỏ (excistion repair) không cần ánh sáng nhờ các nuclease, sau đó thay vào các base đúng. Có thể xảy ra theo nhiều cách: cắt bỏ các base sai hỏng nhờ các enzyme DNA glycosylase. Các enzyme này nhận biết các base bị biến đổi và các điểm mất purine hay mất pyrimidine và thủy giải liên kết N-glycosilic nối base với đường. Rồi enzyme AP endonuclease cắt liên kết đường và phosphate gần base bị biến đổi. Sau đó enzyme thứ ba, deoxyribophosphodiesterase loại bỏ từng nucleotide kế tiếp nhau ở đoạn bị hỏng. Sau đó, DNA polymerase lấp đầy khoảng trống với các nucleotide bổ sung với sợi khuôn còn lại. Enzyme DNA ligase sẽ gắn các khe hở giữa 2 đầu 3′-5′ . Trong tế bào tồn tại một số DNA glycosylase. Chẳng hạn, enzyme uracil- DNA glycosylase cắt uracil khỏi DNA. Uracil tạo thành do đột biến mất nhóm amin ngẫu nhiên ở cytosine, dẫn đến đột biến đồng hoán thay C bằng T. Enzyme này phát hiện ra uracil trên DNA như là một bất thường, chúng sẽ cắt bỏ và sửa sai.

Sửa chữa bằng cắt bỏ không bị hạn chế bởi dimer, cũng có thể sửa chữa các tổn thương khác bằng vật chất hóa học sửa chữa. Xét đối tượng cắt bỏ, sửa chữa bằng cắt bỏ còn có thể chia thành hai loại: sửa chữa cắt bỏ base và sửa chữa loại bỏ nucleotit. Sửa chữa cắt bỏ base loại bỏ trực tiếp base sai hỏng và thay thế nó trong DNA. Ví dụ điển hình là enzyme DNA uracil glicolase, loại các uracil không được bắt cặp với các guanine. Hệ thống sửa chữa cắt bỏ nucleotit cắt bỏ một trình tự bao gồm các base sai hỏng, sau đó 1 đoạn DNA mới được tổng hợp để thay thế nguyên liệu bị cắt bỏ, các hệ thống này phổ biến ở hầu hết các sinh vật.
Chức năng sửa chữa bằng cắt bỏ tồn tại rộng rãi ở sinh vật nhân sơ và sinh vật nhân thực, cũng là phương thức sửa chữa chính ở con người. Động vật gặm nhấm như chuột ngay từ khi sinh ra đã thiếu đi chức năng này.
Năm 1978, học giả Mỹ J.L. Marx phát hiện ra giữa sinh vật nhân thực và sinh vật nhân sơ có cấu tạo chất nhiễm sắc khác nhau, quá trình sửa chữa bằng cắt bỏ cũng khác nhau. Phân tử DNA của sinh vật nhân thực không lộ ra như ở sinh vật nhân sơ, mà là kết cấu nucleosome dạng chuỗi hạt quấn trên histone. Cắt bỏ dimer pyrimidine trong sinh vật nhân thực chia làm hai giai đoạn: cắt nhanh, thời gian 2-3 giờ, chủ yếu cắt bỏ những tổn thương trên bộ phận DNA mà không liên kết với histone; cắt chậm, ít nhất mất 35 giờ, và còn cần loại nhân tử điều khiển nào đó để nhận biết tổn thương này, làm nucleosome ở bộ phận tổn thương trên DNA lộ ra, sau đó phải qua nhiều bước nữa mới hoàn thành việc sửa chữa này, phân tử DNA được sửa chữa quấn quanh histone và hình thành nucleosome mới.
Sửa chữa bằng cách tái tổ hợp
Các vị trí có sự sai hỏng, xảy ra trong một phân tử con, sẽ được phục hồi nhờ sự tái tổ hợp để thu được một bản sao khác của trình tự từ một nguồn không bị sai hỏng. Bản sao này sau đó được dùng để sửa chữa lỗ hổng trên sợi bị hỏng. Khi DNA polymerase gặp thymine dimer hay một số sai lệch khác trên sợi DNA khuôn mẫu, có hai khả năng xảy ra: Polymerase lấp dần lỗ hổng bằng tái bản không theo khuôn mẫu do tái tổ hợp và vượt qua chỗ sai. Polymerase dừng lại và huy động khoảng 1000 nucleotit phía dưới; chỗ trống gián đoạn được lấp đầy với sợi bổ sung từ sợi đôi chị em do tái tổ hợp. Trong cả hai trường hợp, chỗ sai lệch vẫn còn và được loại bỏ sau đó bởi sửa sai trực tiếp hay cắt bỏ.
Sửa chữa hệ thống SOS
Là 1 chức năng trong phản ứng SOS. Trong trường hợp DNA bị hỏng ngừng tái bản, phản ứng SOS sẽ phục hồi tái bản bằng phương thức tái bản “error-prone”. Ở vi khuẩn E. coli người ta đã quan sát thấy sự phá hủy DNA làm mở ra khoảng 20 gen của hệ thống SOS, được điều khiển âm bởi chất ức chế LexA (LexA repressor). Trong trường hợp có nhiều sai hỏng cần cấp cứu, LexA bị kích thích, thay đổi cấu hình, tự cắt và mất hoạt tính ức chế. Lúc đó các gen của hệ thống SOS được mở ra. Nếu quá trình sửa sai thực hiện không kịp thì tế bào phải chấp nhận hoặc bị đột biến hoặc chết. Protein RecA khởi động hệ thống SOS như sau: Hộp SOS là trình tự DNA (operator) dài khoảng 20 bp được nhận biết bởi protein của chất ức chế LexA. Sai hỏng của DNA làm cho RecA khởi động phản ứng SOS, chứa các gen mã hóa cho nhiều enzyme sửa chữa. RecA hoạt hóa hoạt tính tự phân giải tự của LexA. LexA ngăn chặn hệ thống SOS, nhưng phản ứng tự phân giải của nó đã hoạt hóa các gen của hệ thống này.
Khi xảy ra phản ứng SOS, có thể làm tăng chức năng sửa chữa tổn thương. Ví dụ, biểu hiện gen uvr A, uvr B, uvr C, uvr D, ssb, recA, recN và ruv, từ đó tăng cường sửa chữa cắt bỏ, sửa chữa sau sao chép và sửa chữa gẫy chuỗi. còn recA, umuD, umcC lại tham gia cơ chế sửa chữa không rõ rang, làm tăng tỷ lệ tồn tại tế bào, tỷ lệ đột biến cũng tăng lên.
Ngoài tác dụng sửa chữa, phản ứng SOS còn có thể gây ra thay đổi trong phân tách tế bào, giải phóng thể thực khuẩn và hình thức sao chép DNA.
Sửa chữa thích nghi
Năm 1977, học giả Mỹ L. Samson phát hiện thêm một loại phản ứng không giống như sửa chữa SOS ở e.coli, nó có thể sửa chữa methyl hóa ở guanine. Ví dụ, chỉ cần 1mg/ml MNNG nuôi e.coli 2 tiếng, có thể chống lại môi trường e.coli gây ra với nồng độ MNNG cao gấp vài trăm lần. Đây là protein thụ thể methyl được guanine hợp thành trên chuỗi DNA, cysteine và nhóm methyl của protein thụ thể methyl kết hợp thành methylcysteine S, từ đó guanine của methyl hóa được sửa chữa.
Sửa chữa gẫy đoạn
Gồm có sửa chữa gẫy đoạn đơn phân tử DNA, sửa chữa đoạn đôi và sửa chữa nối đoạn nhiễm sắc thể. Dưới tác dụng của men kết nối, các phần gẫy này nhanh chóng được nối lại. Dạng sửa chữa này có hai đặc điểm: một là không ổn định, sau khi nối lại nó vẫn có thể gẫy ra; hai là không chính xác, thường xảy ra lỗi nối.
Sửa chữa kết nối ngang
Bước đầu là tách liên kết ngang dưới tác dụng của glycosylase, bằng việc cắt bỏ codon sửa chữa một chuỗi đơn trong đó; sau đó, dưới tác dụng của endonuclease, sử dụng phương thức sửa chữa bằng cắt bỏ nucleotides, sửa chữa mảnh sợi đơn mặt đối diện.
Nguồn: http://ditruyenhoc.com/cac-dang-ton-thuong-dna-va-phuong-thuc-sua-chua-ton-thuong-dna.html
Related Post
-
 Nông nghiệp thẳng đứng-Arcology
Nông nghiệp thẳng đứng-Arcology
-
 Overview about Molecular Biology in the medical sector
Overview about Molecular Biology in the medical sector
-
 PHÁT HIỆN 2019 NOVEL CORONAVIRUS (2019-nCoV) BẰNG KĨ THUẬT REAL TIME RT-PCR (rRT-PCR)
PHÁT HIỆN 2019 NOVEL CORONAVIRUS (2019-nCoV) BẰNG KĨ THUẬT REAL TIME RT-PCR (rRT-PCR)
-
 ASFV Realtime PCR Kit
ASFV Realtime PCR Kit
-
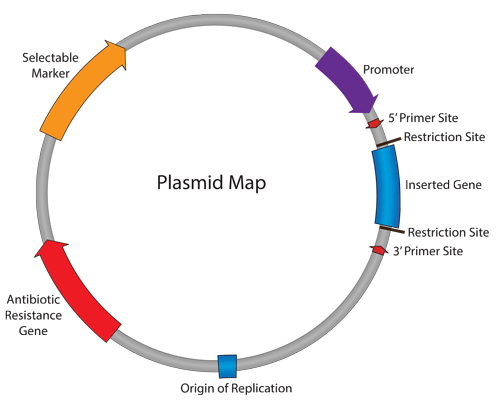 Tái tổ hợp DNA: Khái niệm và phân loại vector
Tái tổ hợp DNA: Khái niệm và phân loại vector
-
 Định lượng miRNA bằng kỹ thuật Real-time RT-PCR trong chẩn đoán sớm ung thư phổi
Định lượng miRNA bằng kỹ thuật Real-time RT-PCR trong chẩn đoán sớm ung thư phổi